In an ideal population
with effective size Ne, the inbreeding
coefficient is simply
F
=1 / (2Ne)
This population then comprises 2Ne
alleles at any locus. Formation
of diploid individuals is a random draw &
replacement exercise from this gene
pool. For any individual, the probability that the first allele
drawn is the same as itself is 1 [think about it]. The probability of drawing the same
allele again is simply the reciprocal of the gene
pool size, thus 1 / (2Ne).
In a finite population with
effective size NeAND some degree of
inbreeding, the inbreeding coefficientF in
the current generation at time t (Ft)
is related to that in the previous generation Ft-1
by a recursion equation,
Ft
= (1/(2Ne)(1)
+ (1 - 1/(2Ne)(Ft-1)
.
The first term applies to inbred individuals,
the second to non-inbred individuals, as follows. The
expectation of drawing any particular allele once is 1 / (2Ne).
As above, if that allele occurs in an inbred individual,
then the expectation that the second allele is identical by
descent with the first is necessarily 1. Otherwise, for non-inbred
individuals, the expectation that the second allele chosen is not
the same asthe first is (1 - 1/(2Ne)).
However, some fraction of the population at time t was inbred in the previous
generation. Call this Ft-1, and the probability of identity by
descent is increased by that factor. The recursion equation then
continues to similar terms for Ft-2, Ft-3,
and so on. Considering the
whole series, the inbreeding coefficient at equilibrium
Feq
is Ft
= Ft-1 = Feq .
[The form of the
recursion equation is identical with that previously used to
estimate the expectations of f(AA)
& f(BB) under inbreeding].
Now consider how mutation changes
the probability of observing two alleles identical by
descent, but not by allelic state. The expectation
that any single allele mutates is µ.
Among individuals with alleles identical by descent, the
expectation that neither allele mutates (and thus
remain identical by descent with each other) is (1 - µ)2.
At equilibrium Feq,
This is an
extremely important equation in evolutionary
population genetics, because given a
constant mutation rate µ,
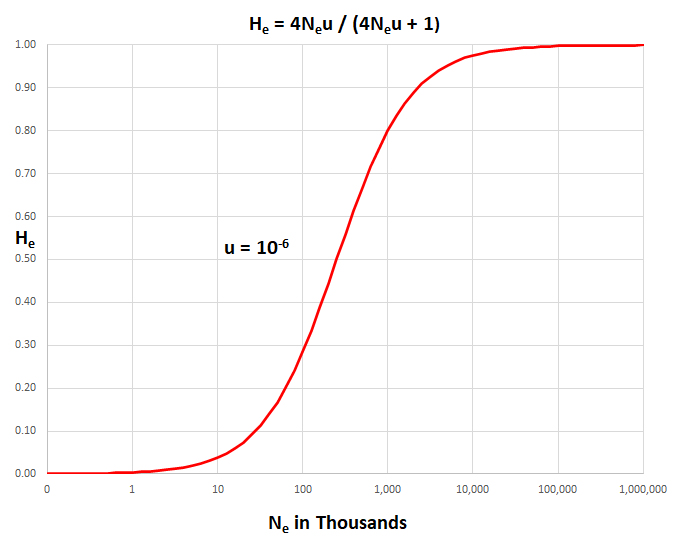
it enables estimation of population size from observed
genetic data. For example, in the semi-log plot
above, for a typical µ
= 10-6, a measured Hobs
= 0.30 predicts Ne
~ 100,000.
HOMEWORK: For µ
= 10-5 and
Hobs = 0.01, 0.02,
0.04, 0.08, 0.16, & 0.32,
calculate the expected Ne.
[HINT: what is the equation for Ne
?]
If Ne
= 1/ µ
, what is the expected value of Hexp?
Does the value of µ
make a difference to the calculation? Why or why not?