The emphasis in this lab is how the pattern of genetic dominance affects the behaviour of alleles subject to negative selection. "Real" examples have been chosen to make the population genetics easier to visualize and discuss; the biology of these examples will be discussed in lecture.
Before coming to lab, read through each scenario and write down the initial conditions of q, W0, W1, and W2 for each part of each exercise. Before running each simulation, check with the instructors to make sure that these values are correct.
NatSel (v5.0) simulates natural selection on a one-locus, two-allele (A & B) model in a monoecious population with random union of gametes (the 'tide pool' model). A seven-digit random number is required when the program is first started. If the first digit is '0', allele frequencies will be called for, otherwise genotype counts will be called for. Required input parameters are the initial allele frequency of B (q = f(B)), population size (N), and the fitness values of the phenotypes corresponding to each of the three genotypes (AA, AB, and BB): W0, W1, and W2, respectively. Input fitness values are normalized to the maximum fitness, which is set as 1.00.
Exactly N individuals are born each generation. Each individual is generated by choosing two alleles at random to form one genotype, where the expectation of a B allele is q. Selection is applied by removal of that individual with an expectation of (1-Wn), where Wn is the normalized fitness of the appropriate phenotype. Genotype proportions and phenotype survival are determined with a pseudo-random number generator, and are therefore not exactly equal to the expected values. The allele frequency q for the next generation is determined from the genotypes of all survivors. Each individual has a life-span of only one generation: it is convenient to think of the parental generation as dying immediately after gamete production.
Output is given in two lines per generation. Allele frequencies, genotype frequencies, and phenotype counts are given both "before" [at birth] and "after" [after selection]. The number of 'selective deaths' is d(N) = Nafter - Nbefore, where N is the sum of phenotype classes. The change in allele frequency in one generation (q) is d(q) = qt - qt-1, the difference in q ('before') between the current and preceding generation. Allele frequency (q) may be displayed graphically at intervals of 10 generations, and are displayed cumulatively for up to 60 generations. Fitness values may be changed every 10 generations.
NatSel is written in FORTRAN77 and compiled with the Microsoft FORTRAN compiler (v.5.0) for PC-DOS machines. It may be copied and distributed freely: the font file HELVB.FON should also be copied and placed in the same directory as the executable program. Please address comments and suggestions by e-mail to "scarr@morgan.ucs.mun.ca"
The phenomenon of industrial melanism in Biston betularia, the common British pepper moth, is perhaps the best studied example of natural selection in the wild. Research on B. betularia has been carried out principally by H. B. D. Kettlewell and co-workers, and is discussed more fully in your text.
Pepper moths are nocturnal animals that during the day remain motionless on the surface of trees. Their major predators are diurnal predators, such as thrushes and other birds, that detect the moths visually and consume them in large numbers. The species is polymorphic for several colour phases. The common form is light grey and speckled, and is highly cryptic (camouflaged) when seen against the bark of the lichen-covered trees common in the rural English countryside. A second colour phase, a melanic (black pigmented) form called "carbonaria", is quite conspicuous on the light-coloured trees of the English countryside. It has been demonstrated experimentally that the two colour phases are subject to differential predation by birds, according to degree of crypsis. The polymorphism has been determined to be under genetic control of a single locus with two alleles. The dominant allele corresponds to the melanic form and the recessive allele to the lighter form.
Prior to the middle of the 19th century, new melanic mutants were rapidly eliminated whenever they arose, and were always rare. The melanic colour phase first began to appear in large numbers in the Manchester area about 1850, and is found in the insect collections of amateur naturalists from that period. This sudden increase coincided with the onset of the Industrial Revolution, which generated extensive air pollution in the form of vast quantities of black soot from the chimneys of coal-burning factories. In heavily industrialized areas (such as Manchester and Birmingham in the southeast of England), forests of previously white, lichen-covered trees became completely blackened. Under these circumstances, the melanic form became more cryptic than the lighter form. By 1900, the proportion of melanic moths in the Manchester area exceeded 90%. Outside of these industrial areas, trees remained relatively uncontaminated and the lighter form continued to be more prevalent. In the latter twentieth century, the death of older trees and pollution control measures, such as installation of "scrubbers" on smokestacks, led to a partial restoration of pre-industrial environmental conditions, so that forests in some previously heavily polluted areas again became "white." The frequency of the melanic form has declined in these areas.
Illustration: Industrial
Melanism in Biston
Start the program by typing "natsel" at the computer prompt. You will be prompted for a seven-digit random number seed [the first digit must be '0']. Then, enter the following values as instructed, in the proper format [confirm with the instructors that the values and format are correct]. Set the initial frequency of the 'light' allele [q = f(B)] as indicated. Set the population size N = 9999. Set the relative fitness values of the three genotypes AA, AB, and BB (W0, W1, W2, respectively) as indicated in each of the scenarios below.
In each of the following five scenarios, record f(B) ('before') at the end of every ten generations. Be prepared to explain to the instructors the changes between the 'before' and 'after' lines in any one generation.
(a) A rural area without pollution, pre-1850. [q = 0.90, 30% selection against the dark phenotype. Continue for up to 60 generations].
(b) A rural area undergoing the post-1850 Industrial Revolution. [q = 0.90, 30% selection against the light phenotype. Continue for 60 generations].
(c) A polluted industrial area, late 19th century. [q = 0.10, 30% selection against the light phenotype. Continue for 60 generations].
(d) An industrial area becoming a "smokeless zone," late 20th century. [q = 0.10, 30% selection against the dark phenotype. Continue for 60 generations].
(e) The British government has plans to establish Whittickshire Forest as a new wildlife preserve, to be located in a former industrial area in the north of England. The plan is to clean up the environment, and then to release a small number of light moths into the (predominantly dark) local population. What is the likely fate of such an experiment? [Re-start NatSel with a new random-number seed (enter '1' as the first digit). Start with _____ homozygous dark moths, zero heterozygous moths, and ______ (homozygous) light moths, and 30% selection against the dark phenotype. Continue for 60 generations]. [Values for blanks will be supplied by the instructors]
On the data sheet, RECORD q = f(B) ('before') and f(BB) at the beginning, and at the end of every ten generations, for each scenario. Use these data to SKETCH a graph of the change in q and the frequency of the light phenotype over 60 generations. Use these data to answer the questions.
Biology 3900 - Principles of Evolution
NAME _________________________
Dr. Carr
Collect the following data in the table in the handout, or a print out of this page of the website
RECORD q = f(B) [frequency of the recessive,
'light' allele] and f(BB) [frequency of the 'light' phenotype] at
the start of each scenario and after 10, 20, 30, 40, 50 & 60 generations.
[Record all values to two decimals.]
| 0 | 10 | 20 | 30 | 40 | 50 | 60 | |
| f(B) / f(BB) | (B) / f(BB) | f(B) / f(BB) | (B) / f(BB) | (B) / f(BB) | (B) / f(BB)) | (B) / f(BB) | |
| a | |||||||
| b | |||||||
| c | |||||||
| d | |||||||
| e |
SKETCH the graph of f(B) and f(BB) you obtained for scenarios a, b, c, & d (not e) (one graph for each scenario). Use these to answer the questions on the answer sheet.
1. How rapidly does strong negative selection modify the frequency of a rare dominant (scenario a) allele as compared with a rare recessive (scenario c) allele? Can either sort of allele ever be completely eliminated from the population by selection? Why is there a difference?
2. How rapidly does strong negative selection modify the frequency of a common recessive (scenario b) allele as compared with a common dominant (scenario d) allele? How quickly does the phenotype of the population change (HINT: how many generations are required for the 'common' phenotype to become 'uncommon')? Why is there a difference?
3. What is the difference in the behavior of a very rare (scenario e) as compared with a rare (scenario d) advantageous phenotype? What happens as the phenotype becomes less rare?
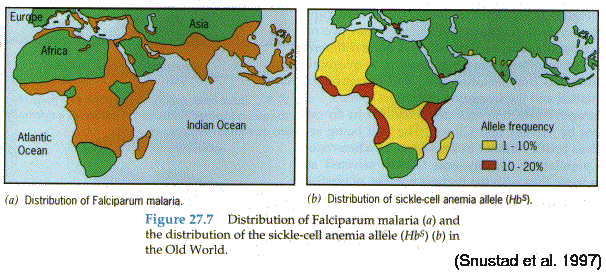
The sickle-cell allele occurs most commonly in human populations from West and Central Africa, in whom it reaches frequencies as high as 16%. These areas are also characterized by high incidence of malaria, and several lines of evidence indicate that the high frequency of this lethal allele is maintained by the increased resistance of AS heterozygotes to malaria. Besides the geographical correlation, (1) the frequency of the sickle-cell trait increases with age among African populations, (2) hospital records indicate increased morbidity due to malaria among AA homozygotes as compared with AS heterozygotes, and (3) clinical tests indicate that the malarial plasmodium parasite is less able to infect AS red cells. In malarial environments, the relative fitness of AA homozygotes seems to be substantially reduced with respect to the AS heterozygotes.
Sickle-cell anemia is a major health problem in the black community of North America, most of whose ancestors originated in West Africa. In the absence of malaria, the heterozygous advantage of the sickle-cell trait is absent, and the S allele is subject to uniform directional selection. A major social concern in the North American black community stems from proposals that prospective parents with the sickle-cell trait ought to obtain prenatal diagnosis and consider abortion of AA (or even AS) fetuses, or alternatively, that they ought to refrain voluntarily from having children.
Illustration: Distribution of
HbS versus malaria in Africa
____________________
Further information on Sickle-Cell Anemia is available locally from:
Canadian Sickle Cell Society
PO Box 13156 / Station A
St. John's NF A1B 4A4
Run NatSel with the following parameters. Let q = f(S), where S is the allele for sickle-cell hemoglobin. Set the population size at N = 9999. The fitness of AA homozygotes is 1.00 in a non-malarial environment, or 0.50 in a malarial environment. In either environment, the fitness of AS heterozygotes is 1.0 and the fitness of SS homozygotes is 0.0.
RECORD the values indicated in the table on the data sheet. The following scenario is continuous over 60 generations, and traces a human population as it moves between different selective environments.
(a) Consider a population of hunter-gatherers from a non-malarial environment in East Africa that initially carries the allele for sickle-cell hemoglobin at a frequency of f(S) = 0.05. [Enter q, N, and W0, W1, and W2 as appropriate]. Continue for 20 generations.
(b) Suppose this population moves westward and begins farming in a high-malarial environment in West Africa. [Continue from part (a): change the fitness values as appropriate, and continue for 20 more generations.]
(c) West African blacks were brought to North America
beginning about 400 years ago. Assume that North America is a non-malarial
environment. [Continue from part (b): change the fitness values as appropriate,
and continue for 20 more generations].
i. f(S) [ = f(B)]
ii. Total deaths [ = d(N)]
iii. Deaths due to sickle-cell [ = # BB]
iv. Deaths due to malaria [ = d(N) - # BB]
v. Mean fitness (W) of the population
[ = (total number of survivors after selection) / 10,000
].
| 0 | 5 | 15 | 25 | 35 | 45 | 55 | |
| (S) [= f(B)] | |||||||
| Total deaths | |||||||
| Sickle-cell deaths | |||||||
| Malaria deaths | |||||||
| Mean Fitness (W) |
SKETCH the graph of f(S). Use the graph and your data to answer the questions on the answer sheet.
0.5+
|
0.4+
|
0.3+
f(S) |
0.2+
|
0.1+
|
0.0+_________|_________|_________|_________|_________|_________|
0
10
20
30 40
50
60
generations
1. In part (b), explain the patterns of deaths due to malaria or sickle-cell anemia after the population moves to the malarial environment (at generation 20).
2. CALCULATE the expected frequency of the S allele between generations 30 & 40 (see your lecture notes or your text). Compare this with the observed frequency in this interval. Explain how f(S) is maintained in this interval.
3. What happens to the mean fitness (W) of the population when the mode of selection changes at 20 generations (hint: what is the total population size after selection in generations 25 and 35)? In a malarial environment, at which value of f(S) is the total population size after selection maximized? Explain.
4. Compare the rate of increase of the frequency of the S allele between generations 20 & 30 with the rate of decrease between generations 40 & 60 (compare the shape of the curves). Why is there a difference (hint: how does the fitness of the AA phenotype change with respect to AS in these two intervals)?
5. In part (c), approximately how many generations are required before f(S) declines to the original value (f(S) = 0.05)? How long would it take to eliminate the S allele if all persons with sickle-cell "trait" (AS individuals) voluntarily choose not to have children? If 50% or 10% of carriers restrained? [FOR FURTHER THOUGHT: What are the social policy and ethical implications of such a solution?]
6. In this model of overdominant selection, selection
against AA in malarial environments has been set unrealistically
high in order to make the principles clearer. If the actual observed frequency
of S in West African populations (q = 0.16) represents the
true equilibrium frequency, calculate the selection coefficient associated
with the A allele.
{kind=link}
{kind=link}